减数分裂是真核生物配子形成过程中一种特殊的细胞分裂方式,是生殖细胞产生的前提。同源染色体之间正确的识别、配对是减数分裂过程中染色体相互作用的开始,对于后续染色体的正确分离至关重要。目前,同源染色体相互精确识别并完成配对的过程和分子机理尚不十分清楚。
中国科学院遗传与发育生物学研究所韩方普研究组长期从事植物减数分裂研究。前期在玉米中观察到减数分裂早前期有同源着丝粒配对的现象,优先于端粒花束的形成,与着丝粒功能直接相关(Zhang et al. 2013. Plant Cell)。近年来在很多物种减数分裂早前期都观察到着丝粒配对或聚集的现象,表明减数分裂早前期的着丝粒配对在大多数真核生物中是普遍存在的现象。着丝粒配对在减数分裂过程中具体是起着什么样的作用,又是如何被精细调控的,还需要进行深入的研究。寻找调控植物减数分裂早前期着丝粒配对的关键因子并探讨其作用机理,将对揭示同源染色体配对的分子机理提供新的认识。
韩方普研究组收集玉米减数分裂I早前期和中期I后的花药,利用染色质免疫共沉淀获得与着丝粒特异组蛋白CENH3 结合的蛋白,通过质谱分析鉴定到减数分裂早前期着丝粒开始配对时与CENH3 结合的候选蛋白为玉米中的SMC3(structure maintenance of chromosome 3)。在有丝分裂过程中,从间期到后期,SMC3定位在染色质上,在染色体臂上和着丝粒区都能检测到SMC3的信号,并且着丝粒区信号富集。减数分裂过程中,从细线期到粗线期,SMC3以点状信号定位在染色质上,并且在着丝粒区有大量富集(图A-C),减数分裂细线期到粗线期是着丝粒配对的时期,SMC3在着丝粒区的大量富集说明SMC3可能参与减数分裂早前期着丝粒的配对。
为了验证SMC3的功能,构建了SMC3基因的CRISPR/Cas9载体和RNAi载体,并进行了玉米的农杆菌转化。CRISPR/Cas9介导的突变率为100%的smc3突变体植株生长极度缓慢,并且在有丝分裂过程中,出现高比例的姐妹染色单体提早分离(图D)或粘连异常(图E),以及染色体异常分离(F)。说明SMC3对维持姐妹染色单体间的粘连至关重要。在SMC3 RNAi转基因植株中,减数分裂细线期到粗线期,着丝粒不配对,着丝粒区SMC3信号减弱,染色质上SMC3信号消失,粗线期染色质形态异常。说明SMC3不仅对维持姐妹染色单体间的粘连至关重要,也参与减数分裂早前期着丝粒配对过程,为揭示减数分裂过程中同源染色体配对的分子机制提供了新的依据。
该论文于2020年1月29日在线发表于The Plant Cell杂志(DOI:10.1105/tpc.19.00834)。韩方普研究组工作人员张晶为第一作者,该研究得到国家自然科学基金的资助。
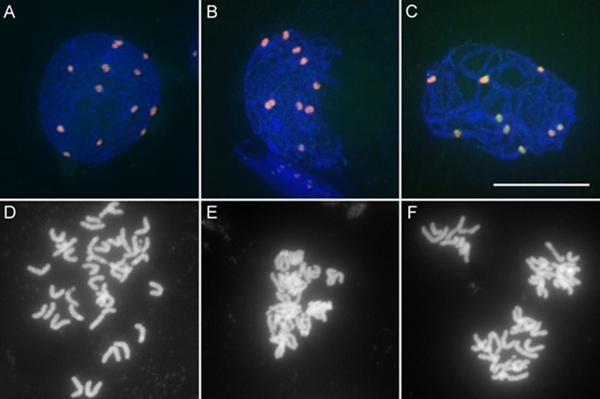
图: SMC3在减数分裂过程中的动态分布和smc3突变体细胞学特征
(A)野生型减数分裂细线期,(B)野生型减数分裂晚偶线期,(C)野生型减数分裂粗线期,
(D)smc3突变体姐妹染色单体提早分离,(E)smc3突变体姐妹染色单体粘连异常,
(F)smc3突变体染色体异常分离。
绿色为SMC3抗体信号,红色为CENH3抗体信号,标尺长度10微米。